Green Fluorescent FAM-FLICA® Caspase-1 (YVAD) Assay Kit

This in vitro assay employs the green fluorescent inhibitor probe FAM-YVAD-FMK to label active caspase-1 enzyme in living cells. Analyze samples using fluorescence microscopy, a fluorescence plate reader, or flow cytometry.


{kind=link}
{kind=link}
{kind=link}
- Prepare samples and controls
- Dilute 10X Apoptosis Wash Buffer 1:10 with diH20.
- Reconstitute FLICA with 50 μL DMSO.
- Dilute FLICA 1:5 by adding 200 μL PBS.
- Add diluted FLICA to each sample at 1:30 (e.g., add 10 μL to 290 μL of cultured cells).
- Incubate approximately 1 hour.
- Remove media and wash cells 3 times: add 1X Apoptosis Wash Buffer and spin cells.
- If desired, label with additional stains, such as Hoechst, Propidium Iodide, 7-AAD, or an antibody.
- If desired, fix cells.
- Analyze with a fluorescence microscope, fluorescence plate reader, or flow cytometer. FAM-FLICA excites at 492 nm and emits at 520 nm.
If working with adherent cells, please see the manual for additional protocols.
Kit 98: 100 Tests
Product Specific References
| PMID | Publication |
| 39999677 | Liu, L, et al. 2025. Microglial pyroptosis induced by SENP7 via the cGAS/STING/IRF3 pathway contributes to neuronal apoptosis. Cytokine, 156893. |
| 39982689 | Wang, C, et al. 2025. Exosomal lncRNA RMRP-shuttled by Olfactory Mucosa-Mesenchymal Stem Cells Suppresses Microglial Pyroptosis to Improve Spinal Cord Injury via EIF4A3/SIRT1. Molecular Neurobiology, . |
| 39934921 | Laanesoo, A, et al. 2025. NLRP1 Is a Prominent Inflammasome Sensor Found in Bronchial Epithelial Cells in Asthma and Can Be Activated by Rhinovirus A16. Clinical and Experimental Allergy, . |
| 39871308 | Chang, Y, et al. 2025. ROS-regulated SUR1-TRPM4 drives persistent activation of NLRP3 inflammasome in microglia after whole-brain radiation. Acta Neuropathologica Communications, 16. |
| 39716284 | Lai, Y, et al. 2024. Novel approach to alleviate lupus nephritis: targeting the NLRP3 inflammasome in CD8+CD69+CD103+ TRM cells. Journal of Translational Medicine, 1139. |
| 39547282 | Leal, VNC, et al. 2024. Bruton's tyrosine kinase (BTK) and matrix metalloproteinase-9 (MMP-9) regulate NLRP3 inflammasome-dependent cytokine and neutrophil extracellular trap responses in primary neutrophils. The Journal of Allergy and Clinical Immunology, . |
| 39736255 | Wāng, Y, et al. 2024. Carbonaceous cores serve as surrogates for environmental particulate matter inducing vascular endothelial inflammation via inflammasome activation. Journal of Hazardous Materials, 137011. |
| 39691768 | Yang, J, et al. 2024. Polydatin-Mediated Inhibition of HSP90α Disrupts NLRP3 Complexes and Alleviates Acute Pancreatitis. Research, 551. |
| 39681234 | Wang, Z, et al. 2024. Dimethyl fumarate alleviates Staphylococcus pseudintermedius-induced cell damage by inhibiting pyroptosis and bacterial virulence. Experimental Eye Research, 110210. |
| 39642726 | Zhang, M., et al. 2024. NLRP3 inflammasome mediates pyroptosis of alveolar macrophages to induce radiation lung injury. Journal of Hazardous Materials, 136740. |
| 39494472 | Long, J., et al. 2024. PD-1+CD8+ T Cell-Mediated Hepatocyte Pyroptosis Promotes Progression of Murine Autoimmune Liver Disease. Advanced Science , e2407284. |
| 39447412 | Pan, C, et al. 2024. Interleukin-38 ameliorates myocardial Ischemia-Reperfusion injury via inhibition of NLRP3 inflammasome activation in fibroblasts through the IL-1R8/SYK axis. International Immunopharmacology, 113428. |
| 39408668 | Camilli, S, et al. 2024. Determining the Feasibility of a Cadmium Exposure Model to Activate the Inflammatory Arm of PANoptosis in Murine Monocytes. International Journal of Molecular Sciences, . |
| 39263574 | Chencen, L., et al. 2024. (+)-catechin protects PC12 cells against CORT-induced oxidative stress and pyroptosis through the pathways of PI3K/AKT and Nrf2/HO-1/NF-κB. Frontiers in Pharmacology, 1450211. |
| 39207402 | Lee, C., et al. 2024. Galectin-9 Mediates the Functions of Microglia in the Hypoxic Brain Tumor Microenvironment. Cancer Research, . |
| 39199694 | Finiuk, N., et al. 2024. The Proapoptotic Action of Pyrrolidinedione-Thiazolidinone Hybrids towards Human Breast Carcinoma Cells Does Not Depend on Their Genotype. Cancers, 0. |
| 39119941 | Mendonça, L.O., et al. 2024. Unexplained fever with consumptive syndrome in the elderly: two cases of VEXAS syndrome with inflammasome dysregulation. Clinical and Experimental Immunology, . |
| 39115441 | Zheng, X., et al. 2024. Eclipta prostrata improves alveolar development of bronchopulmonary dysplasia via suppressing the NLRP3 inflammasome in a DLD-dependent manner. Pediatric Pulmonology, . |
| 38972660 | Li, J, et al. 2024. Mode of cell death in the penile cavernous tissue of type 1 diabetes mellitus rats. The journal of sexual medicine, 652-662. |
| 38941593 | Lucena Lage, S, et al. 2024. Inflammasome activation in patients with Kaposi sarcoma herpesvirus (KSHV)-associated disorders. Blood, 0. |
| 38900640 | Pelletier, AN, et al. 2024. A pre-vaccination immune metabolic interplay determines the protective antibody response to a dengue virus vaccine. Cell reports, 114370. |
| 38864913 | Zhao, LJ, et al. 2024. Heparan sulfate acts as an activator of the NLRP3 inflammasome promoting inflammatory response in the development of acute pancreatitis. Journal of gastroenterology, 0. |
| 38804602 | Guan, X, et al. 2024. Inhibition of HDAC2 sensitises antitumour therapy by promoting NLRP3/GSDMD-mediated pyroptosis in colorectal cancer. Clinical and translational medicine, e1692. |
| 38838402 | Zhan, Z.Y., et al. 2024. NETs contribute to psoriasiform skin inflammation: A novel therapeutic approach targeting IL-36 cytokines by a small molecule tetrahydroxystilbene glucoside. Phytomedicine : international journal of phytotherapy and phytopharmacology, 155783. |
| 38773379 | Xu, X., et al. 2024. BRD4 promotes gouty arthritis through MDM2-mediated PPARγ degradation and pyroptosis. Molecular medicine (Cambridge, Mass.), 67. |
| 38761855 | Rodrigues, F.D.S., et al. 2024. Cannabidiol prevents LPS-induced inflammation by inhibiting the NLRP3 inflammasome and iNOS activity in BV2 microglia cells via CB2 receptors and PPARγ. Neurochemistry international, 105769. |
| 38815927 | Xiao, F., et al. 2024. Dexmedetomidine improves mitophagy and pyroptosis through the ALKBH5/FUNDC1 axis during epidural-related maternal fever. Advances in medical sciences. |
| 38715828 | Huang, Y., et al. 2024. Elabela mitigates the early stage of inflammation in sepsis by inhibiting pyroptosis. American journal of translational research, 1165-1176. |
| 38676641 | Gao, M, et al. 2024. Nano-microflora Interaction Inducing Pulmonary Inflammation by Pyroptosis. Environmental science & technology. |
| 38670314 | Healy, L.D., et al. 2024. An orthosteric/allosteric bivalent peptide agonist comprising covalently linked protease activated receptor-derived peptides mimics in vitro and in vivo activities of activated protein C. Journal of thrombosis and haemostasis : JTH. |
| 38450750 | Wu, F, et al. 2024. Increased platelet-CD8+ T cell aggregates displaying high activation, exhaustion, and tendency to death correlate with disease progression in people with HIV-1. Journal of leukocyte biology. |
| 38169429 | Pinzone, M.R., et al. 2024. Pharmacological approaches to promote cell death of latent HIV reservoirs. Current opinion in HIV and AIDS, 56-61. |
| 38567442 | Chen, X, et al. 2024. Disulfiram Improves Fat Graft Retention by Modulating Macrophage Polarization Via Inhibition of NLRP3 Inflammasome-Mediated Pyroptosis. Aesthetic surgery journal. |
| 38484719 | Zhan, J, et al. 2024. LncRNA ZFAS1 Alleviated NLRP3 Inflammasome-Mediated Pyroptosis through Regulating miR-96-5p/Smad7 Signaling in Allergic Rhinitis. International archives of allergy and immunology, 1-14. |
| 38534380 | Montero-Vega, M.T., et al. 2024. Fluvastatin Converts Human Macrophages into Foam Cells with Increased Inflammatory Response to Inactivated Mycobacterium tuberculosis H37Ra. Cells. |
| 38445369 | Nishimura, K, et al. 2024. Beneficial Effects of Human Schwann Cell-Derived Exosomes in Mitigating Secondary Damage After Penetrating Ballistic-Like Brain Injury. Journal of neurotrauma. |
| 38411635 | Zack, S.R., et al. 2024. Altered vacuole membrane protein 1 (VMP1) expression is associated with increased NLRP3 inflammasome activation and mitochondrial dysfunction. Inflammation research: official journal of the European Histamine Research Society ... [et al.]. |
| 38397063 | Caocci, M., et al. 2024. HIV Infection Drives Foam Cell Formation via NLRP3 Inflammasome Activation. International journal of molecular sciences. |
| 38362146 | Zhang, T., et al. 2024. Sesamin ameliorates nonalcoholic steatohepatitis through inhibiting hepatocyte pyroptosis in vivo and in vitro. Frontiers in pharmacology, 1347274. |
| 38417794 | Sharma, J., et al. 2024. ORMDL3 regulates NLRP3 inflammasome activation by maintaining ER-mitochondria contacts in human macrophages and dictates ulcerative colitis patient outcome. The Journal of biological chemistry, 107120. |
| 38420829 | Liao, L., et al. 2024. TRIM6 Promotes ROS-Mediated Inflammasome Activation and Pyroptosis in Renal Tubular Epithelial Cells via Ubiquitination and Degradation of GPX3 Protein. Frontiers in bioscience (Landmark edition), 58. |
| 38234046 | Tang, R.F., et al. 2024. LncRNA SNHG1 alleviates myocardial ischaemia-reperfusion injury by regulating the miR-137-3p/KLF4/TRPV1 axis. ESC heart failure. |
| 38183100 | Xie, S., et al. 2024. (Pro)renin receptor mediates tubular epithelial cell pyroptosis in diabetic kidney disease via DPP4-JNK pathway. Journal of translational medicine, 26. |
| 38213616 | Horiuchi, Y., et al. 2024. Infection of tumor cells with Salmonella typhimurium mimics immunogenic cell death and elicits tumor-specific immune responses. PNAS nexus, pgad484. |
| 38043264 | Li, Y., et al. 2024. Extracellular histones exacerbate heat stroke-induced liver injury by triggering hepatocyte pyroptosis and liver injury via the TLR9-NLRP3 pathway. International immunopharmacology, 111305. |
| 38127952 | Sabbione, F., et al. 2023. Regulation of human neutrophil IL-1β secretion induced by Escherichia coli O157:H7 responsible for hemolytic uremic syndrome. PLoS pathogens, e1011877. |
| 38089811 | Li, D., et al. 2023. Staphylococcus aureus activates NRLP3-dependent IL-1β secretion from human conjunctival goblet cells using α toxin and toll-like receptors 2 and 1. Frontiers in cellular and infection microbiology, 1265471. |
| 38088496 | Luo, R., et al. 2023. HAGLR, stabilized by m6A modification, triggers PTEN-Akt signaling cascade-mediated RPE cell pyroptosis via sponging miR-106b-5p. Journal of biochemical and molecular toxicology, e23596. |
| 38047519 | Gao, J., et al. 2023. METTL14 knockdown inhibits the pyroptosis in the sepsis-induced acute lung injury through regulating the m6A modification of NLRP3. Experimental lung research, 220-230. |
| 38099557 | Fujita, Y., et al. 2023. C5a stimulation induces caspase-1 activation and mature IL-1β production in human peripheral blood mononuclear cells. Immunological medicine, 1-8. |
| 37945612 | Liu, Y., et al. 2023. Cryo-electron tomography of NLRP3-activated ASC complexes reveals organelle co-localization. Nature communications, 7246. |
| 37966628 | Peng, J., et al. 2023. Neural Stem Cell Extracellular Vesicles Carrying YBX1 Inhibited Neuronal Pyroptosis Through Increasing m6A-modified GPR30 Stability and Expression in Ischemic Stroke. Translational stroke research. |
| 37976936 | Jiang, F., et al. 2023. Inhibition of macrophage pyroptosis ameliorates silica-induced pulmonary fibrosis. Ecotoxicology and environmental safety, 115693. |
| 37924754 | Tong, Z., et al. 2024. Brassisterol A, a new ergosterol from co-cultivation of fungi attenuates neuroinflammation via targeting NLRP3/caspase-1/GSDMD pathway. Bioorganic Chemistry, 106955. |
| 37903858 | Pruenster, M., et al. 2023. E-selectin-mediated rapid NLRP3 inflammasome activation regulates S100A8/S100A9 release from neutrophils via transient gasdermin D pore formation. Nature immunology, . |
| 37929625 | de Pablo, N., et al. 2023. Lipin-2 regulates the antiviral and anti-inflammatory responses to interferon. EMBO reports, e57238. |
| 37798443 | den Hartigh, A.B., et al. 2023. Muscimol inhibits plasma membrane rupture and ninjurin-1 oligomerization during pyroptosis. Communications biology, 1010. |
| 37831404 | Fu, Q., et al. 2023. ACT001 alleviates inflammation and pyroptosis through the PPAR-γ/NF-κB signaling pathway in LPS-induced alveolar macrophages. Genes & genomics, . |
| 37830540 | Williams, J.G., et al. 2023. Streptococcus pyogenes emm98.1 variants activate inflammatory caspases in human neutrophils. Virulence, 2264090. |
| 36280654 | Moraes, C., et al. 2023. Microglial NLRP3 Inflammasome Induces Excitatory Synaptic Loss Through IL-1β-Enriched Microvesicle Release: Implications for Sepsis-Associated Encephalopathy. Molecular Neurobiology, 481-494. |
| 36308669 | Liang, C., et al. 2023. Silencing lncRNA KCNQ1OT1 reduced hepatic ischemia reperfusion injury-induced pyroptosis by regulating miR-142a-3p/HMGB1 axis. Molecular and Cellular Biochemistry, 1293-1305. |
| 36604548 | Chao, YY, et al. 2023. Human TH17 cells engage gasdermin E pores to release IL-1α on NLRP3 inflammasome activation. Nature immunology, 295-308. |
| 36650381 | Mitchell, C.A., et al. 2023. Stromal niche inflammation mediated by IL-1 signalling is a targetable driver of haematopoietic ageing. Nature cell biology, 30-41. |
| 36595704 | Marques-da-Silva, C., et al. 2023. AIM2 sensors mediate immunity to Plasmodium infection in hepatocytes. Proceedings of the National Academy of Sciences of the United States of America, e2210181120. |
| 36640341 | Meyers, A.K., et al. 2023. Pyruvate dehydrogenase kinase supports macrophage NLRP3 inflammasome activation during acute inflammation. Cell reports, 111941. |
| 36736455 | Vaher, H., et al. 2023. Skin colonization with S. aureus can lead to increased NLRP1 inflammasome activation in patients with atopic dermatitis. Journal of Investigative Dermatology, 1268-1278. |
| 36646008 | Hou, L., et al. 2023. Human umbilical cord mesenchymal stem cell-derived extracellular vesicles alleviated silica induced lung inflammation and fibrosis in mice via circPWWP2A/miR-223-3p/NLRP3 axis. Ecotoxicology and environmental safety, 114537. |
| 36863071 | Manna, S., et al. 2023. Nanovaccine that activates the NLRP3 inflammasome enhances tumor specific activation of anti-cancer immunity. Biomaterials, 122062. |
| 36827376 | Reyes, M., et al. 2023. Microscale combinatorial stimulation of human myeloid cells reveals inflammatory priming by viral ligands. Science advances, eade5090. |
| 36508821 | Chen, Y., et al. 2023. Cadmium exposure promotes thyroid pyroptosis and endocrine dysfunction by inhibiting Nrf2/Keap1 signaling. Ecotoxicology and environmental safety, 114376. |
| 36749596 | Ramadan, A., et al. 2023. The NLRP3 Inflammasome Is Required for Protection Against Pseudomonas Keratitis. Investigative ophthalmology & visual science, 11. |
| 36876152 | Cui, Y., et al. 2023. Neutrophil Extracellular Traps Induce Alveolar Macrophage Pyroptosis by Regulating NLRP3 Deubiquitination, Aggravating the Development of Septic Lung Injury. Journal of Inflammation Research, 861-877. |
| 37015177 | Franklin, M., et al. 2023. Activation of inflammasomes and their effects on neuroinflammation at the microelectrode-tissue interface in intracortical implants. Biomaterials, 122102. |
| 36967061 | Sun, L., et al. 2023. Purinergic Receptor P2 × 7 Contributes to Abdominal Aortic Aneurysm Development via Modulating Macrophage Pyroptosis and Inflammation. Translational research : the journal of laboratory and clinical medicine. |
| 36907865 | Hedbrant, A., et al. 2023. Occupational quartz and particle exposure affect systemic levels of inflammatory markers related to inflammasome activation and cardiovascular disease. Environmental health : a global access science source, 25. |
| 37622117 | Bourne, J., et al. 2023. Megakaryocyte NLRP3 hyperactivation induces anemia and potentiates inflammatory response in mice. Frontiers in immunology, 1226196. |
| 36978239 | Li, Q., et al. 2023. Polysaccharide Nanofiber-Stabilized Pickering Emulsion Microparticles Induce Pyroptotic Cell Death in Hepatocytes and Kupffer Cells. Small (Weinheim an der Bergstrasse, Germany), e2207433. |
| 37068092 | Johnson, A.F., et al. 2023. Constitutive secretion of pro-IL-18 allows keratinocytes to initiate inflammation during bacterial infection. PLoS pathogens, e1011321. |
| 37137441 | Karema-Jokinen, V., et al. 2023. Crosstalk of protein clearance, inflammasome, and Ca2+ channels in retinal pigment epithelium derived from age-related macular degeneration patients. The Journal of biological chemistry, 104770. |
| 37167640 | Liu, L., et al. 2023. Parthenolide targets NLRP3 to treat inflammasome-related diseases. International immunopharmacology, 110229. |
| 37000116 | Zhang, Y., et al. 2023. Ubiquitin-specific protease 11 Aggravates Ischemia-reperfusion-induced Cardiomyocyte Pyroptosis and Injury by Promoting TRAF3 Deubiquitination. Balkan medical journal, 205-214. |
| 37284442 | Zhou, W., et al. 2023. Disulfiram with Cu2+ alleviates dextran sulfate sodium-induced ulcerative colitis in mice. Theranostics, 2879-2895. |
| 37300869 | Li, Z., et al. 2023. Cadmium-induced pyroptosis is mediated by PERK/TXNIP/NLRP3 signaling in SH-SY5Y cells. Environmental toxicology. |
| 37265187 | Miao, R.F., et al. 2023. LncRNA CDKN2B-AS1 interacts with LIN28B to exacerbate sepsis-induced acute lung injury by inducing HIF-1α/NLRP3-mediated pyroptosis. The Kaohsiung journal of medical sciences. |
| 37341818 | Xie, Y., et al. 2023. Knockdown of USF2 inhibits pyroptosis of podocytes and attenuates kidney injury in lupus nephritis. Journal of molecular histology. |
| 37429494 | Wang, D., et al. 2023. BDE-47 flame retardant exposure induces microglial pyroptosis and cognitive deficits by activating the mtROS-NLRP3 axis via Sirt3 downregulation and is salvaged by honokiol. Environmental pollution (Barking, Essex : 1987), 122158. |
| 37389784 | Shi, B., et al. 2023. NLRP3 activation in macrophages promotes acute intestinal injury in neonatal necrotizing enterocolitis. World journal of pediatrics : WJP. |
| 37418838 | Xu, K., et al. 2023. Lycium ruthenicum Murr. anthocyanins inhibit hyperproliferation of synovial fibroblasts from rheumatoid patients and the mechanism study powered by network pharmacology. Phytomedicine : international journal of phytotherapy and phytopharmacology, 154949. |
| 37417868 | Kulsuptrakul, J., et al. 2023. A human-specific motif facilitates CARD8 inflammasome activation after HIV-1 infection. eLife. |
| 37457695 | Kim, J.K., et al. 2023. Resistance of hypervirulent Klebsiella pneumoniae to cathepsin B-mediated pyroptosis in murine macrophages. Frontiers in immunology, 1207121. |
| 37459784 | Zhang, M., et al. 2023. MiR-223-3p attenuates radiation-induced inflammatory response and inhibits the activation of NLRP3 inflammasome in macrophages. International immunopharmacology, 110616. |
| 37402829 | Pampuscenko, K., et al. 2023. Extracellular tau stimulates phagocytosis of living neurons by activated microglia via Toll-like 4 receptor-NLRP3 inflammasome-caspase-1 signalling axis. Scientific reports, 10813. |
| 37535431 | Li, J., et al. 2023. Pulmonary Delivery of Specialized Pro-Resolving Mediators-Based Nanotherapeutics Attenuates Pulmonary Fibrosis in Preclinical Animal Models. ACS nano, 15354-15370. |
| 37625726 | Wang, D., et al. 2023. NLRP3 inflammasome-mediated pyroptosis involvement in cadmium exposure-induced cognitive deficits via the Sirt3-mtROS axis. The Science of the total environment, 166478. |
| 37622117 | Bourne, J.H., et al. 2023. Megakaryocyte NLRP3 hyperactivation induces mild anemia and potentiates inflammatory response in mice. Frontiers in immunology, 1226196. |
| 37629052 | Alam, M.A., et al. 2023. Deficiency of Caspase-1 Attenuates HIV-1-Associated Atherogenesis in Mice. International journal of molecular sciences. |
| 37654461 | Chen, A., et al. 2023. Association between NLRP3 inflammasome and periprocedural myocardial injury following elective PCI. Heliyon, 8. |
| 37764917 | Maxson, M., et al. 2023. Mycobacterium tuberculosis Central Metabolism Is Key Regulator of Macrophage Pyroptosis and Host Immunity. Pathogens, 1109. |
| 37748566 | Zhou, Ling, L., et al. 2023. Extracellular ATP (eATP) inhibits the progression of endometriosis and enhances the immune function of macrophages. Biochimica et biophysica acta. Molecular basis of disease, 166895. |
| 37748281 | Wu, Zhimin, Z., et al. 2023. MiR-495-3p attenuates cell pyroptosis and endometritis through inhibiting the activation of NLRP3 inflammasome in bovine. Molecular immunology, 75-85. |
| 37477437 | Dong, Xijie, X., et al. 2023. INTRINSIC/EXTRINSIC APOPTOSIS AND PYROPTOSIS CONTRIBUTE TO THE SELECTIVE DEPLETION OF B CELL SUBSETS IN SEPTIC SHOCK PATIENTS. Shock (Augusta, Ga.), 345-353. |
| 36644875 | Zhou, Z.Y., et al. 2023. Nocardia seriolae mediates liver granulomatous chronic inflammation in Micropterus salmoides through pyroptosis. Journal of fish diseases. |
| 35091064 | Zhang, Y., et al. 2022. HMGB2 causes photoreceptor death via down-regulating Nrf2/HO-1 and up-regulating NF-κB/NLRP3 signaling pathways in light-induced retinal degeneration model. Free radical biology & medicine. |
| 35061085 | Mónaco, A., et al. 2022. Inflammasome activation, NLRP3 engagement and macrophage recruitment to tumor microenvironment are all required for Salmonella antitumor effect. Cancer immunology, immunotherapy:CII. |
| 35167493 | Marek-Iannucci, S., et al. 2022. Targeting IRE1 endoribonuclease activity alleviates cardiovascular lesions in a murine model of Kawasaki disease vasculitis. JCI insight. |
| 35237259 | Wallace, H., et al. 2022. Crosstalk Between Pyroptosis and Apoptosis in Hepatitis C Virus-induced Cell Death. Frontiers in Immunology, 788138. |
| 35124564 | Yao, Y., et al. 2022. Ginsenoside Rd attenuates cerebral ischemia/reperfusion injury by exerting an anti-pyroptotic effect via the miR-139-5p/FoxO1/Keap1/Nrf2 axis. International immunopharmacology, 108582. |
| 35367811 | Liu, Z., et al. 2022. XBP1 deficiency promotes hepatocyte pyroptosis by impairing mitophagy to activate mtDNA-cGAS-STING signaling in macrophages during acute liver injury. Redox Biology, 102305. |
| 35355981 | Lučiūnaitė, A., et al. 2022. Activation of NLRP3 Inflammasome by Virus-Like Particles of Human Polyomaviruses in Macrophages. Frontiers in immunology, 831815. |
| 35246034 | Blake, M.K., et al. 2022. ERAP1 is a critical regulator of inflammasome-mediated proinflammatory and ER stress responses. BMC immunology, 9. |
| 35212950 | Banerjee, I., et al. 2022. In Vitro Assays to Study Inflammasome Activation in Primary Macrophages. Methods in molecular biology (Clifton, N.J.), 45258. |
| 35435672 | Li, Y., et al. 2022. Tumor Microenvironment-Responsive Yolk-Shell NaCl@Virus-Inspired Tetrasulfide-Organosilica for Ion-Interference Therapy via Osmolarity Surge and Oxidative Stress Amplification. ACS nano. |
| 35449370 | Fu, H., et al. 2022. CircACTR2 in macrophages promotes renal fibrosis by activating macrophage inflammation and epithelial-mesenchymal transition of renal tubular epithelial cells. Cellular and molecular life sciences : CMLS, 253. |
| 35462787 | Dai, M., et al. 2022. The Modulation of Interferon Regulatory Factor-1 via Caspase-1-Mediated Alveolar Macrophage Pyroptosis in Ventilator-Induced Lung Injury. Mediators of inflammation, 1002582. |
| 35433045 | Neudorf, H., et al. 2022. Examining the Effect of Consuming C8 Medium-Chain Triglyceride Oil for 14 Days on Markers of NLRP3 Activation in Healthy Humans. Journal of nutrition and metabolism, 7672759. |
| 35654037 | Panicker, N., et al. 2022. Neuronal NLRP3 is a parkin substrate that drives neurodegeneration in Parkinson's disease. Neuron. |
| 35788117 | McLemore, A.F., et al. 2022. Somatic gene mutations expose cytoplasmic DNA to co-opt the cGAS-STING-NLRP3 axis in Myelodysplastic syndromes. JCI insight. |
| 35842415 | Trugilho, M.R.O., et al. 2022. Platelet proteome reveals features of cell death, antiviral response and viral replication in covid-19. Cell death discovery, 324. |
| 36463191 | Cai, Z., et al. 2022. Plumbagin rescues the granulosa cell's pyroptosis by reducing WTAP-mediated N6-methylation in polycystic ovary syndrome. Journal of Ovarian Research, 126. |
| 35945940 | Yin, H., et al. 2022. HMGB1-activatied NLRP3 inflammasome induces thrombocytopenia in heatstroke rat. PeerJ, e13799. |
| 36063959 | Sun, Y., et al. 2022. A single-beam of light priming the immune responses and boosting cancer photoimmunotherapy. Journal of controlled release : official journal of the Controlled Release Society, 734-747. |
| 36288704 | Jiang, Y., et al. 2022. Gasdermin D restricts anti-tumor immunity during PD-L1 checkpoint blockade. Cell reports, 111553. |
| 36252682 | Yuan, X., et al. 2022. Endothelial Acid Sphingomyelinase Promotes NLRP3 Inflammasome and Neointima Formation during Hypercholesterolemia. Journal of lipid research, 100298. |
| 36279668 | Hu, R., et al. 2022. Edaravone dexborneol provides neuroprotective benefits by suppressing NLRP3 inflammasome-induced microglial pyroptosis in experimental ischemic stroke. International immunopharmacology, 109315. |
| 36252479 | Wan, Y., et al. 2022. Ursolic acid alleviates Kupffer cells pyroptosis in liver fibrosis by the NOX2/NLRP3 inflammasome signaling pathway. International immunopharmacology, 109321. |
| 36224321 | Wen-Yuan, W., et al. 2022. mTORC1-Dependent and GSDMD-Mediated Pyroptosis in Developmental Sevoflurane Neurotoxicity. Molecular neurobiology. |
| 36379253 | Liu, W., et al. 2022. Spirodalesol analog 8A inhibits NLRP3 inflammasome activation and attenuates inflammatory disease by directly targeting adaptor protein ASC. The Journal of biological chemistry, 102696. |
| 36538076 | Liu, F., et al. 2022. Mitophagy-promoting miR-138-5p promoter demethylation inhibits pyroptosis in sepsis-associated acute lung injury. Inflammation research : official journal of the European Histamine Research Society ... [et al.], . |
| 36514346 | Claude, L., et al. 2022. Platelet caspase-1 and Bruton tyrosine kinase activation in patients with COVID-19 is associated with disease severity and reversed in vitro by ibrutinib. Research and practice in thrombosis and haemostasis, e12811. |
| 33432150 | Dufies, O., et al. 2021. Escherichia coli Rho GTPase-activating toxin CNF1 mediates NLRP3 inflammasome activation via p21-activated kinases-1/2 during bacteraemia in mice. Nature microbiology. |
| 33430344 | Mutala, L.B., et al. 2021. The Caspase-1/IL-18 Axis of the Inflammasome in Tumor Cells: A Modulator of the Th1/Tc1 Response of Tumor-Infiltrating T Lymphocytes in Colorectal Cancer. Cancers. |
| 33262522 | Baroja-Mazo, A., et al. 2021. Author Correction: Early endosome autoantigen 1 regulates IL-1β release upon caspase-1 activation independently of gasdermin D membrane permeabilization. Scientific reports, 21356. |
| 33459112 | Wang, Z., et al. 2021. LncRNA MIAT downregulates IL-1β, TNF-ɑ to suppress macrophage inflammation but is suppressed by ATP-induced NLRP3 inflammasome activation. Cell cycle (Georgetown, Tex.), 44936. |
| 33405023 | Cao, X., et al. 2021. CHRFAM7A Overexpression Attenuates Cerebral Ischemia-Reperfusion Injury via Inhibiting Microglia Pyroptosis Mediated by the NLRP3/Caspase-1 pathway. Inflammation. |
| 33449124 | Andrade-Santos, J.L., et al. 2021. Differences in pyroptosis of recent thymic emigrants CD4+ T Lymphocytes in ART-treated HIV-positive patients are influenced by sex. Immunogenetics. |
| 33430331 | Bhattarai, N., et al. 2021. Effects of Resvega on Inflammasome Activation in Conjunction with Dysfunctional Intracellular Clearance in Retinal Pigment Epithelial (RPE) Cells. Antioxidants. |
| 33620070 | Ranéia E Silva, P.A., et al. 2021. Inflammatory effect of Bothropstoxin-I from the Bothrops jararacussu venom mediated by NLRP3 inflammasome involves ATP and P2X7 receptor. Clinical science. |
| 35095880 | Lage, S.L., et al. 2021. Persistent Oxidative Stress and Inflammasome Activation in CD14highCD16- Monocytes From COVID-19 Patients. Frontiers in immunology, 799558. |
| 33357072 | Delaney, M.A., et al. 2020. Avoidance of the NLRP3 Inflammasome by the Stealth Pathogen, Coxiella burnetii. Veterinary pathology, 300985820981369. |
Question: What is the difference between YVAD and WEHD and when is it an advantage using one or the other?
Answer: Our new product offering FAM-WEHD-FMK is similar to our existing FAM-YVAD-FMK Assay. Both of these peptide sequences are known to target caspase 1,4, and 5. The WEHD sequence is thought to be a “better” caspase-1 target, as the kcat/kM rate is higher for WEHD vs YVAD (meaning faster conversion of substrate product by the enzyme). However, please note that if our understanding of how FLICA works is correct, the FLICA probe never actually binds to the enzyme via the YVAD or WEHD sequence, but rather the FMK moiety, then perhaps these faster binding kinetics are something of a moot point. In practice the performance characteristics of the two product are very similar. In our lab they were shown to be virtually indistinguishable. Nevertheless, we decided to carry both options so that customers can select their preferred targeting sequence based on their individual needs and experience.
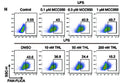
Question: Customer is not seeing a difference between control and induced cells(induction with LPS+ATP). Can we help with optimization? Parameters: macrophages induced from THP-1 cells, using 50 ng/ml PMA for 48 hr Cells in 12 well plates at 3×10^5 cells/well Three groups: experimental with HIV, Positive Control and Untreated. Given fresh media 24 hrs then added 1 ug/ml LPS for 24 hr then 5 mM ATP for 2 hr
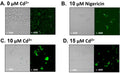
Answer: In our lab, we actually saw a greater response in the THP-1 monocytes (not PMA-primed), we had the greatest response with LPS exposure at 100 ng/mL + 5 mM ATP for 24 hours. In our THP I monocyte studies we found induction levels ranging from 10-30% (average was 26.2%) in 24 hour (LPS/ATP exposure) samples compared to 3-8% in negative controls. When working with THP-1 cells primed with PMA to become macrophage-like, in general we were able to achieve better results with lower LPS concentrations and exposure periods than with the THP-1 monocytes. For instance, exposure to 10ng/mL LPS for 2 hours without any supplemental ATP was sufficient to produce the desired effect. I am a bit concerned that the customer’s use of 1 ug/mL LPS for 24 hours may be too high concentration/exposure period and the susceptible cells are moving through pyroptosis, lysing, and are lost from the positive control sample well prior to even receiving the FAM-YVAD-FMK stain. If this is the case, they are missing the period when more of the positive control cells would be stain positive with FAM-FLICA. I would encourage them to experiment with lower LPS concentrations and exposure periods and see if their results are improved. It is also important to note caspase-1 is rapidly secreted by macrophages after its activation by the inflammasome pathway. Therefore, it turns out macrophages might not be the best cell model for use with this product. We have also been working with nigericin, as an alternate inducing agent.
Question: The component FAM-YVAD-FMK Part#665 vial in the kit is empty. Please help me to solve this problem.
Answer: All of our FLICA products, including FAM-YVAD-FMK, are lyophilized as part of the manufacturing process. The vials contain such a small amount of material (µg quantities) that the green FAM-FLICA reagents are nearly invisible in the amber vials. It may be visible as a slight iridescent sheen on the sides of the vial. Per the instructions in our manual, the FLICA vials are reconstituted in DMSO and diluted into PBS and subsequently diluted into cell culture media for staining cells. In order to check that the FLICA vial contains the proper lyophilized reagent, please check the appearance of the DMSO-reconstituted FLICA reagent. It should be orange in appearance and once diluted 1:5 in PBS, the FAM-FLICA reagent should be yellow in appearance.