Green Fluorescent FAM-FLICA® Caspase-3/7 (DEVD) Assay Kit
This in vitro assay employs the green fluorescent inhibitor probe FAM-DEVD-FMK to label active caspase 3 and caspase 7 enzymes in living cells. Analyze samples using fluorescence microscopy, a fluorescence plate reader, or flow cytometry.

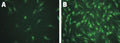


Figure 1. Cell death in primary rat hippocampal neurons. (A) is a composite of FLICA® FAM-DEVD-FMK (B), PI (C), and Hoechst (D). DNA is labeled in 4 cells (D), 3 cells are caspase-positive (B), 2 are membrane-compromised (C). One cell is in early apoptosis (green but not red). Two FLICA®-positive cells are also PI-positive (red, C) (becoming membrane compromised) and are in the late stages of apoptosis rather than necrosis. Data courtesy of Dr. Z. Kahraman Akozer, University of Maryland.
Click on image to zoom
{kind=link}
{kind=link}
Ships: 1-2 business days
- Prepare samples and controls
- Dilute 10X Apoptosis Wash Buffer 1:10 with diH20.
- Reconstitute FLICA with 50 µL DMSO.
- Dilute FLICA 1:5 by adding 200 µL PBS.
- Add diluted FLICA to each sample at 1:30 (e.g., add 10 µL to 290 µL of cultured cells).
- Incubate approximately 1 hour.
- Remove media and wash cells 3 times: add 1X Apoptosis Wash Buffer and spin cells.
- If desired, label with additional stains, such as Hoechst, Propidium Iodide, 7-AAD, or an antibody.
- If desired, fix cells.
- Analyze with a fluorescence microscope, fluorescence plate reader, or flow cytometer. FAM-FLICA excites at 492 nm and emits at 520 nm.
If working with adherent cells, please see the manual for additional protocols.
Kit 93: 25 Tests
FLICA Caspase-3/7 Reagent (FAM-DEVD-FMK), 1 vial, #653
10X Apoptosis Wash Buffer, 15 mL, #635
Fixative, 6 mL, #636
Propidium Iodide, 1 mL, #638
Hoechst 33342, 1 mL, #639
Kit Manual
Kit 94: 100 Tests
FLICA Caspase-3/7 Reagent (FAM-DEVD-FMK), 4 vials, #653
10X Apoptosis Wash Buffer, 60 mL, #634
Fixative, 6 mL, #636
Propidium Iodide, 1 mL, #638
Hoechst 33342, 1 mL, #639
Kit Manual
Product Specific References
| PMID | Publication |
| 39613751 | Gudneppanavar, R., et al. 2024. Ezrin drives adaptation of monocytes to the inflamed lung microenvironment. Cell Death & Disease, 864. |
| 39427254 | Jorge-Cruz, C.Y., et al. 2024. Guinea pig spermatozoa adhesion to an immobilized fibronectin matrix alters their physiology and increases their survival. Biology of Reproduction, . |
| 39366751 | Xia, Y, et al. 2024. PARP inhibitors enhance antitumor immune responses by triggering pyroptosis via TNF-caspase 8-GSDMD/E axis in ovarian cancer. Journal for Immunotherapy of Cancer, . |
| 39335166 | Chia, W.T., et al. 2024. Okanin Inhibits Cell Growth and Induces Apoptosis and Pyroptosis in Oral Cancer. Cancers, . |
| 39199694 | Finiuk, N., et al. 2024. The Proapoptotic Action of Pyrrolidinedione-Thiazolidinone Hybrids towards Human Breast Carcinoma Cells Does Not Depend on Their Genotype. Cancers, . |
| 38972660 | Li, J, et al. 2024. Mode of cell death in the penile cavernous tissue of type 1 diabetes mellitus rats. The journal of sexual medicine, 652-662. |
| 38788367 | Radomska, D., et al. 2024. Evaluation of anticancer activity of novel platinum(II) bis(thiosemicarbazone) complex against breast cancer. Bioorganic chemistry, 107486. |
| 37834182 | Scopelliti, F., et al. 2023. Functional TRPA1 Channels Regulate CD56dimCD16+ NK Cell Cytotoxicity against Tumor Cells. International journal of molecular sciences, . |
| 37001390 | Ivasechko, I., et al. 2023. Molecular design, synthesis and anticancer activity of new thiopyrano[2,3-d]thiazoles based on 5-hydroxy-1,4-naphthoquinone (juglone). European journal of medicinal chemistry, 115304. |
| 37128606 | Capitini, C., et al. 2023. APP and Bace1: Differential effect of cholesterol enrichment on processing and plasma membrane mobility. iScience, 106611. |
| 37200190 | Fike, A.J., et al. 2023. STAT3 signaling in B cells controls germinal center zone organization and recycling. Cell reports, 112512. |
| 37247053 | Wankell, M., et al. 2023. Testing Cell Migration, Invasion, Proliferation, and Apoptosis in Hepatic Stellate Cells. Methods in molecular biology (Clifton, N.J.), 43-54. |
| 37371034 | Rok, J., et al. 2023. The Assessment of Anti-Melanoma Potential of Tigecycline-Cellular and Molecular Studies of Cell Proliferation, Apoptosis and Autophagy on Amelanotic and Melanotic Melanoma Cells. Cells, . |
| 37553330 | Matsui, Y., et al. 2023. SNIP1 and PRC2 coordinate cell fates of neural progenitors during brain development. Nature communications, 4754. |
| 37748566 | Zhou, Ling, L., et al. 2023. Extracellular ATP (eATP) inhibits the progression of endometriosis and enhances the immune function of macrophages. Biochimica et biophysica acta. Molecular basis of disease, 166895. |
| 37477437 | Dong, Xijie, X., et al. 2023. INTRINSIC/EXTRINSIC APOPTOSIS AND PYROPTOSIS CONTRIBUTE TO THE SELECTIVE DEPLETION OF B CELL SUBSETS IN SEPTIC SHOCK PATIENTS. Shock (Augusta, Ga.), 345-353. |
| 37760225 | Pezo, Felipe, F., et al. 2023. Slow Freezing of Preserved Boar Sperm: Comparison of Conventional and Automated Techniques on Post-Thaw Functional Quality by a New Combination of Sperm Function Tests. Animals: an open access journal from MDPI, . |
| 35055021 | Rok, J., et al. 2022. The Anticancer Potential of Doxycycline and Minocycline-A Comparative Study on Amelanotic Melanoma Cell Lines. International journal of molecular sciences, . |
| 34994998 | Chiu, Y.J., et al. 2022. Curcumin suppresses cell proliferation and triggers apoptosis in vemurafenib-resistant melanoma cells by downregulating the EGFR signaling pathway. Environmental toxicology, . |
| 35111247 | Huang, C.F., et al. 2022. Quercetin induces tongue squamous cell carcinoma cell apoptosis via the JNK activation-regulated ERK/GSK-3α/β-mediated mitochondria-dependent apoptotic signaling pathway. Oncology letters, 78. |
| 35270004 | González-Sarrías, A., et al. 2022. Milk-Derived Exosomes as Nanocarriers to Deliver Curcumin and Resveratrol in Breast Tissue and Enhance Their Anticancer Activity. International journal of molecular sciences, . |
| 35241512 | Tsai, Y.F., et al. 2022. Gadodiamide Induced Autophagy and Apoptosis in Human Keratinocytes. In vivo (Athens, Greece), 603-609. |
| 35842415 | Trugilho, M.R.O., et al. 2022. Platelet proteome reveals features of cell death, antiviral response and viral replication in covid-20. Cell death discovery, 324. |
| 35939511 | Kawashima, A., et al. 2022. Genome-wide screening identified SEC61A1 as an essential factor for mycolactone-dependent apoptosis in human premonocytic THP-1 cells. PLoS neglected tropical diseases, e0010672. |
| 35967457 | Su, M., et al. 2022. Gasdermin D-dependent platelet pyroptosis exacerbates NET formation and inflammation in severe sepsis. Nature cardiovascular research, 732-747. |
| 36063959 | Sun, Y., et al. 2022. A single-beam of light priming the immune responses and boosting cancer photoimmunotherapy. Journal of controlled release : official journal of the Controlled Release Society, 734-747. |
| 36296570 | Janowska, S., et al. 2022. Synthesis and Anticancer Activity of 1,3,4-Thiadiazoles with 3-Methoxyphenyl Substituent. Molecules, . |